CAROTIPO
Representación gráfica, numérica y morfológica del equipamiento cromosómico de una célula, que es una constante de cada especie. El cariotipo humano comprende 23 pares de cromosomas ordenados en ocho grupos según su aspecto. Toda anomalía cualitativa o cuantitativa de este esquema se traduce en afecciones diversas, generalmente graves. cariotipo masculino normal.
ANOMALÍAS EN CARIOTIPOS
SÍNDROME DE DOWN
El síndrome de Down (SD) es un trastorno genético causado por la presencia de una copia extra  del cromosoma 21 (o una parte del mismo), en vez de los dos habituales (trisomía del par 21), caracterizado por la presencia de un grado variable de retraso mental y unos rasgos físicos peculiares que le dan un aspecto reconocible. Es la causa más frecuente de discapacidad psíquica congénita1 y debe su nombre a John Langdon Haydon Down que fue el primero en describir esta alteración genética en 1866, aunque nunca llegó a descubrir las causas que la producían. En julio de 1958 un joven investigador llamado Jérôme Lejeune descubrió que el síndrome es una alteración en el mencionado par de cromosomas.
del cromosoma 21 (o una parte del mismo), en vez de los dos habituales (trisomía del par 21), caracterizado por la presencia de un grado variable de retraso mental y unos rasgos físicos peculiares que le dan un aspecto reconocible. Es la causa más frecuente de discapacidad psíquica congénita1 y debe su nombre a John Langdon Haydon Down que fue el primero en describir esta alteración genética en 1866, aunque nunca llegó a descubrir las causas que la producían. En julio de 1958 un joven investigador llamado Jérôme Lejeune descubrió que el síndrome es una alteración en el mencionado par de cromosomas.
SINDROME DE TURNER
El síndrome Turner, síndrome Ullrich-Turner o monosomía X es una enfermedad genética caracterizada por la presencia de un solo cromosoma X. Genotípicamente son mujeres (por ausencia de cromosoma Y). A las mujeres con síndrome de Turner les falta parte o todo un cromosoma X. En algunos casos se produce mosaicismo, es decir que la falta de cromosoma X no afecta a todas las células del cuerpo. cariotipo de una mujer con síndrome de turner
SÍNDROME DE EDWARDS
El síndrome de Edwards, también conocido como trisomía 18, es una aneuploidía humana que se caracteriza usualmente por la presencia de un cromosoma adicional completo en el par 18. También se puede presentar por la presencia parcial del cromosoma 18 (trasladación desbalanceada) o por mosaicismo en las células fetales
SINDROME DE PATAU
El síndrome de Patau, trisomía en el par 13 o trisomía D es una enfermedad genética que resulta de la presencia de un cromosoma 13 suplementario. El cariotipo da 47 cromosomas y sirve de diagnóstico prenatal por amniocentesis o cordiocentesis sobre todo si los padres optan por el aborto. Se trata de la trisomía menos frecuente.



















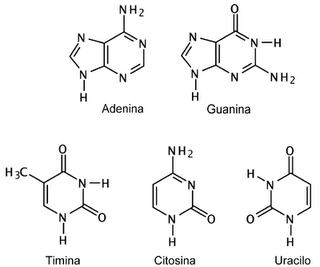
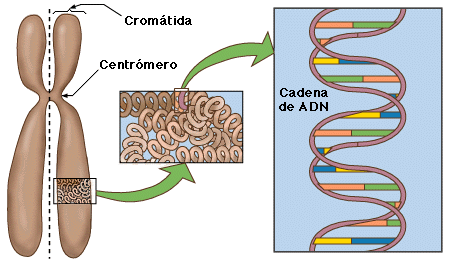

{kind=link}
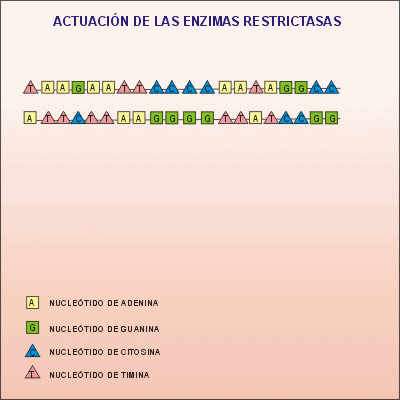{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}