{kind=link}
 ACIDO DESOXIRIBONUCLEICO:ADN
ACIDO DESOXIRIBONUCLEICO:ADNADN es la abreviatura del ácido desoxirribonucleico. Constituye el material genético de los organismos. Es el componente químico primario de los cromosomas y el material del que los genes están formados. En las bacterias y otros organismos unicelulares, el ADN está disribuido por la célula. En organismos más complejos tal como plantas, anímales y otros organismos multicelulares, la mayoria del ADN reside en el núcleo celular. El ADN es un ácido nucleico formado por nucleótidos. Cada nucleótido consta de tres elementos:
- un azúcar: desoxirribosa en este caso (en el caso de ARN o ácido ribonucleico, el azúcar que lo forma es una ribosa),
- un grupo fosfato
- una base nitrogenada
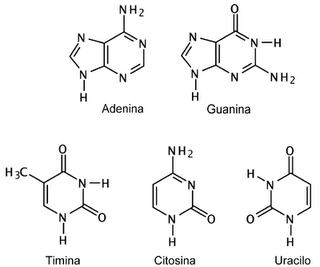
Las bases nitrogenadas que constituyen parte del ADN son: adenina (A), guanina (G), citosina (C) y timina (T). Estas forman puentes de hidrógeno entre ellas, respetando una estricta complementariedad: A sólo se aparea con T (y viceversa) mediante dos puentes de hidrógeno, y G sólo con C (y viceversa)
 mediante 3 puentes de hidrógeno.
mediante 3 puentes de hidrógeno.Los extremos de cada una de las hebras del ADN son denominados 5’-P (fosfato) y 3’–OH (hidroxilo) en la desoxirribosa. Las dos cadenas se alinean en forma paralela, pero en direcciones inversas (una en sentido 5’ → 3’ y la complementaria en el sentido inverso), pues la interacción entre las dos cadenas está determinada por los puentes de hidrógeno entre sus bases nitrogenadas. Se dice, entonces, que las cadenas son antiparalelas.
CROMOSOMAS
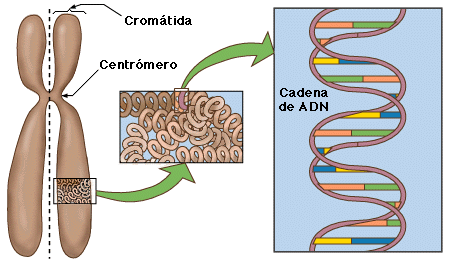
Los cromosomas son las estructuras físicas de la célula eucariota que portan los genes. Estos cromosomas solo son visibles durante la división celular. Mostrando a plenitud sus características morfológicas durante la meta fase.
La dotación cromática humana es de 23 pares, los cuales se clasifican en 22 pares de autosomas y un par de cromosomas sexuales o gonosomas (XX en la mujer y XY en el hombre). Los miembros de cada par son semejantes y se denominan homólogos.
Desde el punto de vista de su composición los cromosomas están formados de DNA y proteínas, donde principalmente con las proteínas básicas llamadas histonas, se forma la llamada fibra de cromatina. Esta cromatina se encuentra de manera des-compactada cuando la célula se encuentra en interfase, razón por la cual los cromosomas no están visibles; pero al momento del comienzo de la profase de la división celular esta fibra va sufriendo un superenrrollamiento que va progresivamente estructurando las dos cromáticas que forman un cromosoma mitótico metafísico.
GENES
Un gen es un segmento corto de ADN, que le dice al cuerpo cómo producir una proteína específica. Hay aproximadamente 30.000 genes en cada célula del cuerpo humano y la combinación de todos los genes constituye el material hereditario para el cuerpo humano y sus funciones.
La composición genética de una persona se llama genotipo.
Los genes están localizados en hebras de ADN, de manera similar a una sarta de cuentas. Las hebras de ADN conforman los cromosomas.
Los cromosomas son pares apareados de una copia de un gen específico. El gen se encuentra en la misma posición en cada cromosoma.

Recientemente, sin embargo, los nuevos descubrimientos han alterado radicalmente esta visión, obligándonos a adoptar una definición bastante más vaga. De acuerdo con ello, un gen es una secuencia de DNA genómico o de RNA que es esencial para especificar una determinada función. Para llevar a cabo su función el gen no necesita ser traducido a proteína, y a veces ni siquiera necesita ser transcrito.
Genes codificadores de proteínas
Eucariotas
Un gen eucariota standard codificador de proteína tiene la estructura que se muestra en la Figura 3 (Li & Graur, pp. 7)
En el flanco 5' se sitúan los promotores (región promotora):
TATA box (19-27 bases antes del punto de comienzo de la transcripción).
CAAT box (situada aún más arriba).
Una o más copias de la caja GC, que rodean la caja CAAT.
Las cajas CAAT y GC controlan la unión inicial de la RNA-polimerasa, mientras que la caja TATA controla el punto de comienzo de la transcripción.
En el flanco 3' se situan los terminadores de transcripción y el sitio de poliadenilación (cola de poly-A).
No es posible definir exactamente donde comienza y donde acaba un gen, es decir, donde empieza el flanco 5' y donde acaba el 3'.
La transcripción comienza en el sitio de inicio de la transcripción (el sitio cap en el transcrito de RNA) y acaba en el sitio de terminación de la transcripción (terminador) que puede coincidir o no con el sitio de poliadenilación. Es decir, que a veces la transcripción puede terminar mucho más abajo del sitio de poliadenilación.
El transcrito primario (pre-mRNA) contiene regiones 5' y 3' que no se van a traducir, exones e intrones (estos últimos tampoco se van a traducir, se eliminan en el proceso de splicing o de maduración del RNA). Todas las secuencias genómicas que permanecen en el RNA maduro después del splicing se consideran exones. Los exones, o partes de exones, que se traducen forman la llamada región codificadora.
Hay varios tipos de intrones, según el mecanismo que se utilice para eliminarlos del pre-mRNA. Aquí nos referiremos a los intrones de genes nucleares que son transcritos por la RNA-polimerasa II y que se cortan enzimáticamente en el proceso de maduración del RNA.
Los sitios de unión (splicing) de intrones y exones son el sitio donante (5') y el sitio aceptor (3'). La inmensa mayoría de los intrones nucleares de eucariotas comienzan con GT y acaban con AG (regla GT-AG).
El número de intrones varía mucho de un gen a otro. Algunos genes tienen varias docenas de intrones, algunos de los cuales pueden alcanzar una longitud de varias Kb. Otros genes, como los de las histonas, no tienen intrones.
La mayor parte de los genes eucarióticos corresponde a los intrones (Figura 4, pp 8 de Li & Graur).
Eubacterias
No contienen intrones.
Los promotores de eubacterias contienen una secuencia -10 y otra -35, así llamadas porque se sitúan ese número de bases antes del punto de inicio de la transcripción. La secuencia -10 contiene el motivo TATAAT o alguna variante, y la secuencia -35 el motivo TTGACA o alguna variante.
Genes de RNA
Su estructura es similar en eucariotas y procariotas.
Generalmente no contienen intrones, aunque en algunos ciliados, hongos y bacterias los hay.
Los transcritos primarios de estos genes suelen sufrir modificaciones posteriores: incorporación de nucleótidos standard y no-standard, modificación de algunos nucleótidos standard en otros no-standard, adición enzimática de secuencias terminales de ribonucleótidos, etc.
Genes reguladores
Genes replicadores (orígenes de replicación): Determinan los sitios de inicio y terminación de la replicación
Genes recombinadores (sitios de recombinación): Sitios de reconocimiento para las enzimas que intervienen en la recombinación.
Genes segregadores: Sitios de unión de los cromosomas a la maquinaria de segregación durante mitosis y meiosis.
Sitios de unión en general para proteínas, hormonas y otras moléculas.
Enhancers (exaltadores): Regulación de la especificidad tisular.
Telómeros: Varios centenares de pares de bases formados por secuencias repetitivas cortas en los extremos de los cromosomas eucarióticos. Ej: CCCTAA en humanos y CCCTAAA en Arabidopsis. El telómero sirve para 'sellar' el extremo del cromosoma, de forma que no sea 'pegajoso' (que no se una a otros cromosomas) y que no sea susceptible al ataque de las exonucleasas. La replicación de los telómeros la lleva a cabo una enzima específica, la telomerasa, que actua como una transferasa terminal. La ausencia se esta enzima en las células puede repercutir en la viabilidad de las mismas, relacionándose con procesos de envejecimiento y muerte celular. Su actividad extemporánea, por el contrario, puede llevar a inmortalidad celular en procesos cancerosos.
No hay comentarios:
Publicar un comentario